什么是腓骨肌萎缩症?
腓骨肌萎缩症是一种罕见的遗传性疾病。
▪ 该病表现为下肢末端(脚和小腿)的肌肉力量下降以及肌肉萎缩,以及后期手和前臂的肌肉力量下降和肌肉萎缩,并且脚和手产生平衡障碍且敏感性降低(触觉、热感和冷感等)通常也与其相关。
▪ 这些表现是由于腿部和手臂神经受损造成的。
▪ 该病大部分起于童年时期或年轻成年时期,症状为走路困难、脚部畸形。该病最初症状出现的年龄、疾病严重程度以及疾病发展个体差异性很大,包括同一家族内的不同成员。
▪ 医疗保健的主要目的在于防止产生并发症, 尤其是肌肉和关节部位(足部护理、拐杖、平衡障碍肌体训练,甚至是骨科手术)的并发症。定期进行适合的运动能力练习可以保持肌肉力量和耐力,并降低患有 CMT 病人的疲劳和疼痛感。
腓骨肌萎缩症的不同形式
腓骨肌萎缩症有超过 60 种的表现形式,按照 3 个标准来进行分类:
- 周围神经受损状态,按神经传导的速度来确定:
• 轴突型:神经传导速度 > 40 m/s,这种速度属于正常或轻度减慢(神经冲动轻度(幅度)减弱),
• 脱髓鞘 (或髓鞘)型:神经传导速度 < 35 m/s,
• 中间型:神经传导速度在 25 m/s 到 45 m/s 之间;
- 遗传传播方式:
常染色体显性遗传
常染色体隐性遗传
伴 X 染色体遗传
- 相关的基因异常
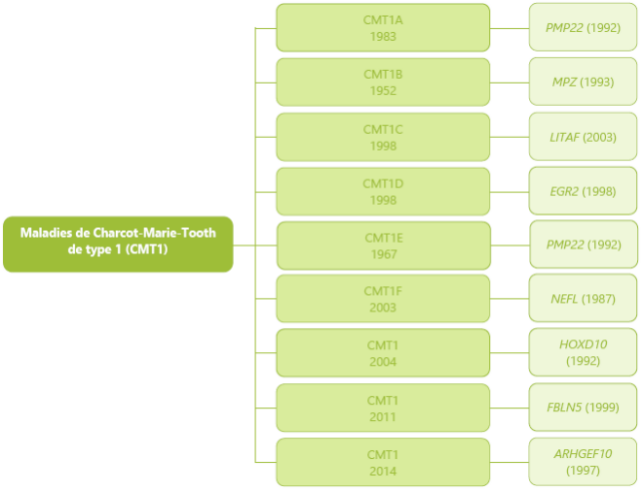
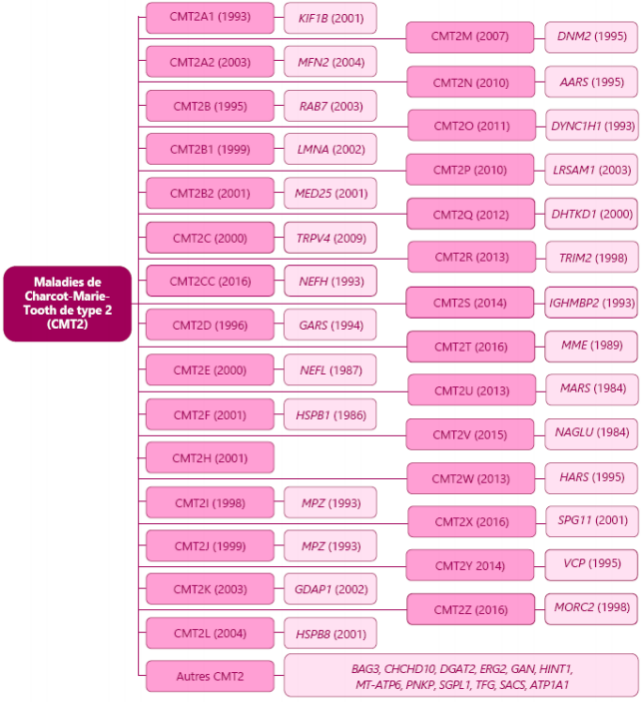
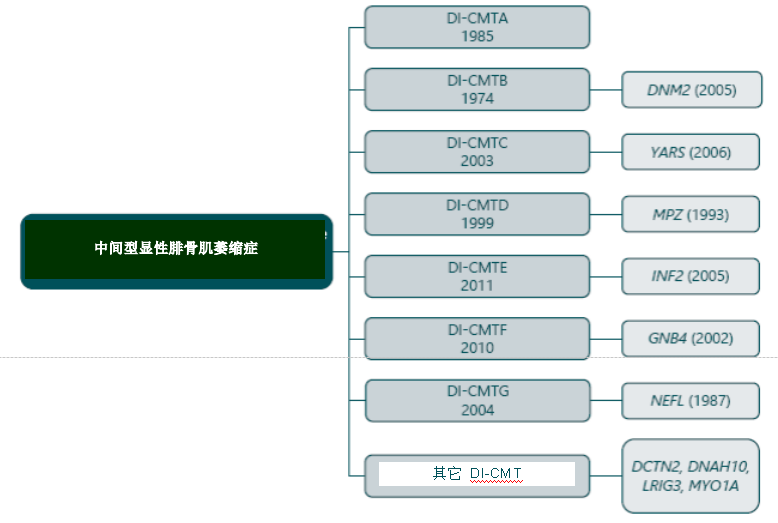
CMT分为 6 大类型:CMT1,CMT2,CMT422,CMTX DI-CMT 和 RI-CMT。
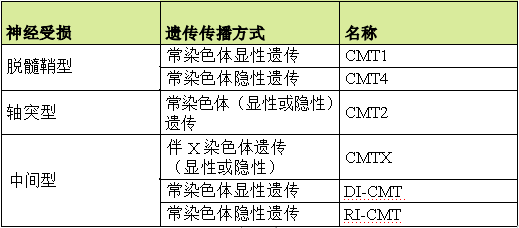
这六种主要类型被分为几个子类(1A 、1B 、1C、2A 、2B),每个都与一个基因异常相对应。 比如,CMT1A 对应的是常染色体显性脱髓鞘型,与 PMP22 基因有关, 而 CMT1B 对应于常染色体显性脱髓鞘型,与 MPZ 基因有关。 这种复杂的分类主要针对于研究 CMT 的医生和研究者,使其可以了解 CMT 中遇到的基因多样性。
腓骨肌萎缩症由什么原因引起?
腓骨肌萎缩症(CMT)是由于 DNA 发生变体导致的疾病,这是一种遗传性疾病。这种 DNA 异常是随机出现的。传递神经冲动较差的周围神经CMT 内涉及的基因异常对周围神经的机能产生了干扰。
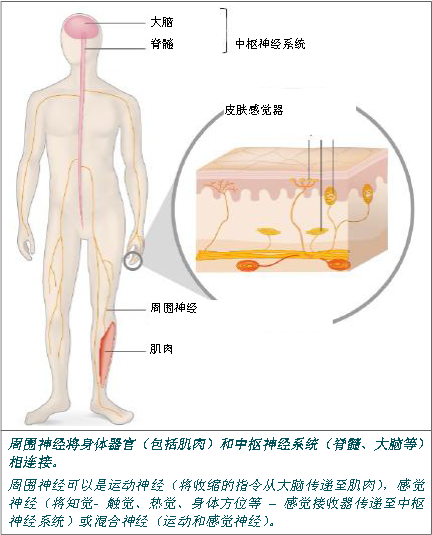
周围神经由神经纤维或轴突构成。轴突传递一种电质信号:神经冲动。许多轴突被独立鞘包围,髓鞘可以更快地传输信号。
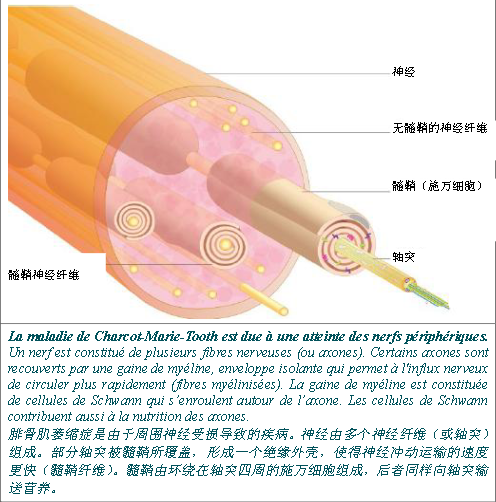
产生髓鞘的是施万细胞,后者确保轴突产生电气绝缘,从而允许神经冲动快速传导。鞘瘤细胞同样参与轴突的生长和营养组成、神经元生存以及神经损伤修复等。
神经传导速度
神经冲动沿神经的速度(或 VCN)是在实施肌电图(或 ENMG)的过程中测量的。
传导速度取决于神经纤维直径以及是否存在髓鞘。正常情况下,传导速度大于40m/s。
肌电图可以对不同形式的 CMT 进行区分:
▪ 脱髓鞘型(或髓鞘型),其神经传导速度低于 35m/s。它是由于基因异常改变了髓鞘或制造髓鞘的施万细胞而导致的。
▪ 轴突型,其神经传导速度大于 40m/s。轴突型 CMT 是由于基因异常导致轴突变形而引起的。越少的轴突参与到神经内进行神经冲动传输,信号也就更弱(我们称之信号幅度更弱)。
▪中间型,该型的神经传导速度在 25 m/s < VCN <45 m/s 之间。
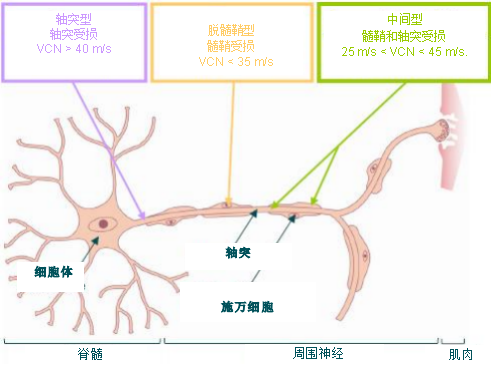
腓骨肌萎缩症是由于连接脊髓和肌肉的周围神经受损导致的疾病。
神经传导速度(VCN)的测定可以将腓骨肌萎缩症的轴突型、脱髓鞘型以及中间型进行区分。
神经越长,越有可能受损
浅表感觉可使我们和外界进行接触:
传递触觉讯息(压力知觉,刺痛知觉)、温度信息或是痛感。
深层感觉主要传递源于肌肉、关节裹(包括关节的部分)、肌腱、韧带的信息,告诉我们身体在空间内的不同部位(本体感受)。
我们正是因为有这种知觉,可以在闭上双眼的情况下让一只手抓住另一只手或者站立保持平衡。
无论信号是否变得更弱或更慢,周围神经传递其讯息的效率都更低。
该讯息有可能是触觉(浅表感觉)的感觉讯息,是空间内平衡需要的身体方位讯息(深层感觉),更或是到达肌肉实施动作的收缩指令(肌肉力量)。
在腓骨肌萎缩症(CMT)中,周围神经受损主要造成四肢末端的感觉障碍以及肌肉刺激缺陷。我们说的是远端肌肉受损。肌肉刺激不足往往会使体积减小(肌肉萎缩)、失去力量(肌肉无力)以及肌肉缩短(肌肉收缩)。远端肌肉受损远端肌肉指的是离身体轴线较远的肌肉,位于四肢端部:上肢即手和前臂的肌肉,下肢则是脚和小腿的肌肉。

四肢远端
不同作用的蛋白质
近几年内,对不同 CMT 型涉及基因编码的蛋白质作用的了解有所增强。
CMT 中改变的主要细胞功能
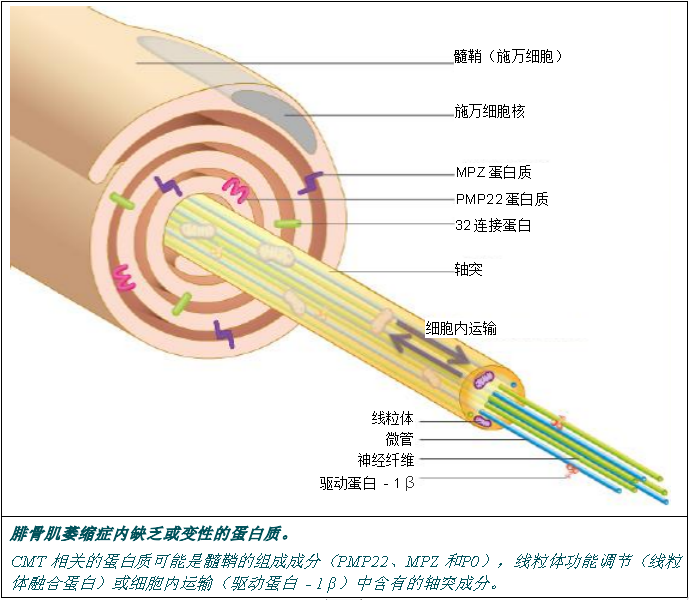
腓骨肌萎缩症中的部分变异基因对参与髓鞘组成、细胞内运输(提供“物质”)的轴突功能、(提供能量) 线粒体功能以及蛋白质合成的蛋白质进行编码。
髓鞘组成

髓鞘由脂质和特定蛋白质构成,在神经元轴突周围形成一层鞘来保护大量的神经纤维,并且可以提高神经冲动的传导速度。
细胞内的运输

细胞内运输指的是整套的运输机制,可以使细胞将物质从一个细胞隔间运输到另一个细胞隔间。在神经细胞中,物质会连续运动,从神经元的细胞体移动至轴突末梢,并反之进行移动。
线粒体的作用

线粒体会产生我们细胞工作不可或缺的能量。轴突含有大量线粒体,来为神经细胞提供其运行所需要的能量。
蛋白质合成

蛋白质合成可以使细胞产生所有细胞运行所需的蛋白质。
髓鞘蛋白质 髓鞘由施万细胞构成,后者围绕在轴突四周形成包裹层。这些包裹层堆积起来就是绝缘鞘(髓鞘),使神经冲动能够快速进行传播。 一些髓鞘组成蛋白质,如 PMP22、P0 蛋白或 32 连接蛋白,都参与形成了腓骨肌萎缩症的不同型式。 P0 蛋白质 MPZ 基因(0 蛋白质髓鞘)编码的 P0 蛋白质是髓鞘中含量最多的蛋白质,因为该蛋白质占髓鞘蛋白质的50%。P0 蛋白质CMT1B、CMT2I、CMT2J 以及 DI-CMTD 几种型式中显示异常。 P0 蛋白质是一种使髓鞘紧凑且稳定不可缺少的蛋白质,因为该蛋白质在施万细胞形成的两层髓鞘中构成了一个连接。 PMP22 蛋白质 周围髓鞘蛋白质 22 或 PMP22 由 PMP22 基因进行编码,其占髓鞘蛋白质的 2-5% 。PMP22 蛋白质的确切作用尚不得而知,但研究提出该蛋白质参与了施万细胞的成长和分化。
PMP22 蛋白主要在周围神经系统内,但我们在中枢神经系统(大脑)中也发现了少量该蛋白。
32 连接蛋白 连接蛋白 32 是一种仅在髓鞘中发现的蛋白质,通过 GJB1 (也称为Cx32)基因编码。当此种蛋白存在异常时,会导致 CMTX1 型疾病。 32 连接蛋白两两组合在一起,形成两个髓鞘层之间的通道以让一层髓鞘层分子传递到另一层髓鞘层上去。 在大多数的情况下,相关的基因异常对通道功能会产生影响,对髓鞘造成破坏并减缓神经冲动的传播速度。 轴突蛋白 部分轴突蛋白参与形成了 CMT 的一种或另一种型式: ▪ 对线粒体功能调节起作用的蛋白(线粒体融合蛋白,GDAP1 蛋白等), ▪ 参与细胞内运输的蛋白(如发动蛋白 2,驱动蛋白- 1 β等), ▪ 其余起不同作用的蛋白(如己糖激酶1,参与到葡萄糖转化为能量的酶等)。 神经细胞线粒体动力学中的蛋白 线粒体是细胞能量中枢,确保产生细胞直接可用的大部分能量。细胞中的线粒体数量根据细胞所需能量是可变的。 线粒体经历持续的合并和分裂(分裂为两个线粒体),来形成新的线粒体,淘汰变性的线粒体并在它们之间进行物质和信息交换。因此,该合并/分裂机制对于线粒体生长、其分布情况以及线粒体网良好状态维持非常重要。 ▪ 在 CMT 中,神经元线粒体的合并/分裂动力学异常是导致神经冲动传递不良的原因,而且可能会对神经元的存活状态进行改变。 ▪ 在对线粒体动力学起作用的蛋白质当中,(CMT2A 中改变的)融合蛋白通过在两个线粒体间形成连接使线粒体相互结合,从而在合并过程中发挥着重要作用。 ▪(CMT2H, CMT2K 和 CMT4A 中改变的) GDAP1 蛋白在线粒体分裂中发挥着重要作用,但其具体作用尚不清楚。 神经细胞胞内运输的蛋白质 胞内运输指的是所有使物质(蛋白质、气泡和细胞单元等)在细胞内部进行循环的机制。 在神经元中,胞内运输对于轴突的生长、改造以及功能维持具有重要重要。 胞内运输同样对髓鞘发展的过程很有必要,它向施万细胞提供髓鞘成分。 参与到 CMT 不同型式的蛋白在神经细胞胞内运输中发挥着重要作用。 ▪ 发动蛋白 2(CMT2M,DI-CMTB)调节参与细胞内囊泡运动微管的稳定性。 ▪ 驱动蛋白-1β(CMT2A1)可以在轴突中沿微管来运输线粒体。 ▪ LITAF (CMT1C) 蛋白质通常都存在于囊泡中参与胞内运输(核内体、溶酶体等)。当这种蛋白质发生异常时, 它会被进行转移并出现在线粒体中。 蛋白合成中的蛋白质 在蛋白质的合成中,氨基酸一个接一个的进行组合来生产蛋白质。氨酰-ARNt 合成酶是酶,可以将氨基酸附着在相应传递的 ADN 上。CMT 中,有的氨酰-ARNt 合成酶是处于异常状态的: - 甘氨酰-ARNt 合成酶(GARS), - 丙氨酰-ARNt 合成酶(AARS), - 甲硫氨酰基-ARNt 合成酶(MARS), - 组氨酰基-ARNt 合成酶(HARS), - 酪氨酰-ARNt 合成酶(YARS), - 赖氨酰-ARNt 合成酶(KARS),
涉及的大量基因
CMT 疾病涉及的基因是逐渐被发现的。
PMP22 基因
1992 年发现了CMT 中涉及的第一个基因:它是(位于17号染色体上的)PMP22 基因, 对 PMP22 进行编码。
▪ CMT1A 型与 PMP22 基因的复制有关: 存在一个 PMP22 基因样本,因此会大量复制 PMP22 蛋白,并减缓神经冲动的传播速度。
▪ 另外两种疾病型式与 PMP22 基因有关:
- CMT1E 型式由于 PMP22 基因突变造成的,同样减缓了神经冲动的传导速度。
- 遗传性压力敏感性神经病变(HNPP)或腊肠样神经病变是由于 PMP22 基因缺失造成的(也就是说至少存在一个 PMP22 基因样本)。因此 PMP22 蛋白产量减少,接着使髓鞘变得脆弱,神经便对持续时间长的机制压迫变得敏感。

中枢神经系统和周围神经系统
之间的差别是什么?
中枢神经系统包括脑部(大脑、小脑和脑干)及其延伸部分和脊髓,受骨结构保护(颅骨盒保护大脑,脊柱保护脊髓)。中枢神经系统分析感官信息、处理动作信息并将收缩指令传递给肌肉。
周围神经系统延长了中枢神经系统,并与之进行密切配合。它是由所有的周围神经组成, 运动神经或感觉神经, 这些神经将中枢神经系统和机体其它神经相连接(器官、肌肉、感觉接收器等)。运动神经把收缩指令传递给肌肉,感觉神经则将感觉-触觉、热觉、身体方位等传递至中枢神经系统。
1 型腓骨肌萎缩症(CMT1) CMT1 是该疾病的脱髓鞘型式,以常染色体显性模式进行遗传。
4 型腓骨肌萎缩症(CMT4) CMT4 是该疾病的脱髓鞘型式,以常染色体隐性模式进行遗传。


2 型腓骨肌萎缩症(CMT2) 该型是轴突型,CMT2 通过常染色体显性或隐性传播。

腓骨肌萎缩症的中间型 CMT 中间型同时表现出髓鞘和轴突都受损的情况。该型是以常染色体显性(DI-CMT)或隐性(RI-CMT)来遗传的。
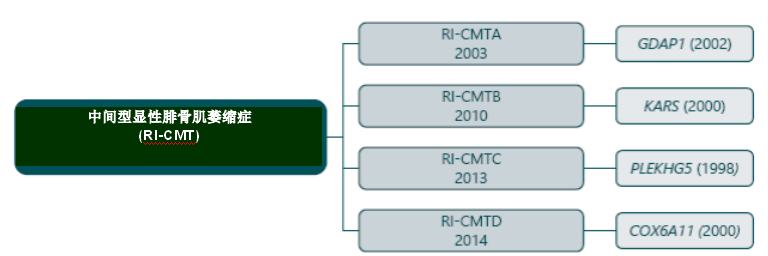
X 伴性腓骨肌萎缩症(CMTX) CMT的X伴性型同时表现出髓鞘和轴突都受损,其传播通常是与X伴性显性遗传相关,极少数情况下为X伴性隐性传播。
腓骨肌萎缩症的临床研究是什么 ?
CMT 的临床研究可以使我们更好地了解疾病的表现方式和疾病的演变方式(疾病自然史),以及采取治疗对疾病症状产生的影响(观察性研究)。临床研究的另一个方面可以评估疾病的潜在治疗效果(候选药物,医学设备等等):这些都是临床试验。
数据库
通过开发患者数据库,可以对患有疾病的人进行统计(如有登记,会进行详细统计)、明确该疾病的自然史,并建立基因型/表现型关系。数据库同样可以对同质的患者人群实施研究, 并为临床研究或试验提供患者招募的便利。
CMT 各种型式中所含基因可以更好地予以识别,但仍然很难准确地确定每项基因异常的临床演变特点或建立基因型/表现型的关系。事实上,除了 CMT1A 是 CMT 最常见的型式以外,大多数其他型式的患者数量都很少。因此,随着时间推移需要建立患者数据库来收集更多 CMT 患者的医学科学数据。
腓骨肌萎缩症的治疗途径有哪些?
由于腓骨肌萎缩症中涉及大量的基因以及疾病中的临床异质性,腓骨肌萎缩症的治疗因此十分复杂。
在 CMT1、CMT2F 和 CMTX1 中,腓骨肌萎缩症的治疗途径以细胞模型和动物模型为基础进行研究(临床前研究)。在 CMT1A,即 CMT 最常见的形态中,则在细胞和动物模型(临床前研究)以及人体(临床研究)上进行研究。
什么是临床前研究?
临床前研究的研究内容是候选药物在培养细胞(活体外)和动物模型(活体内)中的性能。这是在人体中施用候选药物的前提条件。
在候选药物开发的临床前阶段,研究人员会研究分子的药理、药物代谢动力和毒理:作用机理、物理化学特性、化合物在机体内的变异、靶器官、毒性等。通过临床前研究因此可以实现人体服用剂量无毒性作用的初步估计。
这些数据对于向监管机构申请未来药物产品的投放市场许可(AMM)是不可或缺的。
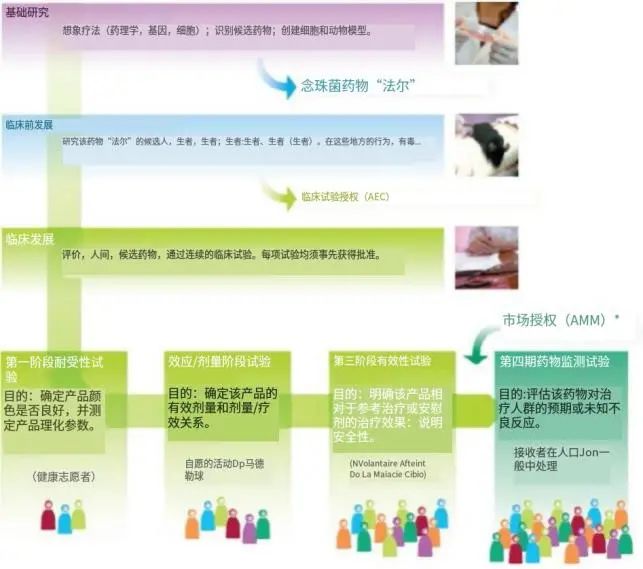
药物的开发步骤
药物的开发从治疗概念(药理治疗、细胞治疗或基因治疗)形成到用于疾病须经过数年(甚至数十年)。
腓骨肌萎缩症的治疗该如何组织?
参与腓骨肌萎缩症(CMT)研究的团队集中在发达国家,尤其是美国、 法国、英国和意大利,同时也有日本、德国、西班牙和澳大利亚。
比利时的情况(更具体地说是安特卫普的研究人员) 较为不同, 因为多年来, 比利时在 CMT 的基因作图领域一直非常投入,但这种疾病的发病率并不比其他地都方要高。
国际会议
▪ 参与腓骨肌萎缩症研究的人员和临床医生会定期参与特别研讨会(讲习班),由欧洲神经肌肉中心(ENMC)组织。这些研讨会汇集了一定数量的参与者(约二十人),能够在合作项目方面进行密集和富有成效的交流。
15 年来,共组织了关于 CMT 的 ENMC 研讨会 6 次,具体涉及以下内容:
- 1A 型腓骨肌萎缩症(2005 年 4 月),
- 评估标准和 CMT 的临床试验(2009 年 9 月),
- CMT 动物模型的临床前研究(2011 年 5 月),
- 与线粒体融合/裂变缺陷有关的神经肌肉疾病;分子机制和治疗策略 (2013 年 4 月),
- 最后一次ENMC研讨会名为《CMT 中的足部手术》,于 2016 年6月举行.这是一次总结用于治疗 CMT 患者足部变形的足部手术不同技术的会议。讨论内容还包括须接受足部手术的患者的最佳手术时间、手术类型和后续方案。
▪ 首届专门讨论 CMT2A 的国际大会于 2014年4月在米兰(意大利)举行,其目的是介绍 CMT2A 研究的最新进展, 并促进潜在的国际科学合作。
▪ 周围神经学会的双年大会还包括一次关于 CMT 的会议。最近一次会议于2018年7月25日在美国巴尔的摩举行。
▪ “国际腓骨肌萎缩症及相关神经病理学大会”会定期汇集临床医生和专门研究这一领域的研究人员。
第6次大会于2016年9月8至10日在威尼斯举行。此次大会上, 向科学界提出了一种新的 CMT 形态分类,以 3 种参数为基础。
- CMT形态的传递模式:常染色体显性形态(AD)、常染色体隐性形态(AR)、伴 X 形态(XL)、偶发形态(Spo); - 神经损伤:脱髓鞘损伤为“De”,轴突损伤为“Ax”,中间损伤 为“In”; - 相关基因名称(尚未知的基因标为“未知”)。 因此,CMT1A 应为“AD-CMT(De)-PMP22”。 为了解科学界对这一分类建议的看法,向来自世界各地的300 名 CMT 专家发出了一份调查问卷。在 107 份回复中,有 65%认为有必要简化目前的分类。 如果新的分类方法被科学界认可,其还必须在专家小组中进行讨论和工作。 腓骨肌萎缩症协会每年还会为 CMT 的相关人员组织一至两次会议,以使其获得关于 CMT 研究和管理的最新信息。
如何参与研究?
在目前的知识状态下,研究人员主要需要从腓骨肌萎缩症患者身上提取的脱氧核糖核酸、细胞和肌肉及神经模型,以继续研究这些疾病中所涉及的各种机制。皮肤碎片也同样有用。
另一种参与研究的方式是参与调查或者临床试验。脱氧核糖核酸(通过采集血液)、皮肤、肌肉捐赠研究小组使用这些采集模型来研究疾病的机制或测试新的治疗途径。

















